|
|
|
Для зеленых автотрофных растений свет является одним из важнейших факторов жизни, поскольку представляет им необходимую лучистую энергию
для фотосинтеза, т.е. участвует в образовании органических веществ, необходимых для роста и развития. Кроме того, свет оказывает непосредственное влияние на рост, на многие
процессы дифференциации в клетках и тканях, на само органообразование. Для жизни растений важно, что в процессе фотосинтеза они продуцируют больше веществ, чем необходимо
для покрытия расходов на дыхание, т. е. образуется положительный баланс веществ, без которого немыслим рост и существование растения. Как и при каких условиях образуется
положительный баланс веществ, эта проблема подлежит экологическому исследованию. Практиков сельского или лесного хозяйства интересует урожай, т. е. продуктивность самого
фотосинтеза. А эколог должен изучить и понять причины различной продуктивности фитоценозов (в связи с различной интенсивностью света) в разных условиях. Кроме того, весьма
важен вопрос, как распределяются ассимиляты, как они используются самим растением и в фитоценозе в целом, т. е. как свет влияет на продуктивность растительного покрова.
В противоположность теплу и воде свет распределен более или менее равномерно, т. е. на Земле фактически нет такой зоны, где бы рост растения не был возможен из-за недостатка
света. Если в полярных областях, где господствует длительная ночь, растения отсутствуют вовсе или их рост очень затруднен, то это связано не с недостатком света, а в первую
очередь с неблагоприятными температурными условиями. Поэтому для расчленения растительности на зоны и подзоны свет играет подчиненную роль. Но его значение особенно велико
в распределении растений на малых площадях, в местообитаниях, т. е. в определении структуры сообщества. Когда мы сравниваем флоры солнечного и теневого местообитаний, то их
различия вызываются в первую очередь условиями освещения, хотя тепловой и водный режимы играют здесь тоже немаловажную роль.
СВЕТОВАЯ ЭНЕРГИЯ И ЕДИНИЦЫ ЕЕ ИЗМЕРЕНИЯ
На верхней границе атмосферы Земля получает почти всю (90 %) радиационную энергию от Солнца. Длина световых волн, достигающих атмосферы, варьирует от 200 до 4000 нм. Единицами
измерения длины световых волн в международной системе единиц (СИ) являются микрометр (мкм) и нанометр (нм): 1мкм = 10−3мм = 10−4см
= 10³нм; 1нм = 10−6мм = 10−7см = 10 ангстремам (А) = 10−9метра.
Огромное количество лучистой энергии Солнца уходит за пределы солнечной системы. И только одна двухмиллиардная часть, т. е. 2 х 10−9лучистой энергии нашего
светила, пройдя более 150 млн. км, доходит до поверхности атмосферы Земли. Это так называемая «солнечная постоянная». Однако не все лучи Солнца достигают поверхности самой Земли.
На высоте около 50км в атмосфере имеется слой озона, который непроницаем для лучей с длиной волн менее 295 нм. Поверхности Земли достигают ультрафиолетовые коротковолновые
(< 400 нм) лучи, составляющие около 10% суммарной радиации; видимые лучи с длиной волн 400 - 750 нм - около 45%; инфракрасная, или «ближняя инфракрасная» (БИКР), с длиной волн
750-4000 нм тоже около 45%. Радиация с длинами волн больше 4000 нм составляет длинноволновую, или дальнюю инфракрасную; в ней в основном происходит тепловое излучение Земли.
Видимая часть спектра (свет как таковой) расположена в границах от 400 до 750 нм или, при большей чувствительности зрения, - от 380-780 им. По обе стороны от этой видимой части
простираются невидимые лучи - инфракрасные (тепловые) и ультрафиолетовые («химические»). Из физиологии известно, что абсорбционная область хлорофилла почти точно совпадает с
абсорбционной областью зрительного пурпура нашего глаза, поэтому для фотосинтеза важен практически лишь видимый свет.
Интервал лучей с длиной волн от 380 до 710 нм (или по другим данным - 400 - 700 нм) принято считать фотосинтетически активной частью радиации (ФАР). ФАР - это такая радиация, квант
которой, поглощаясь молекулой хлорофилла, приводит ее в возбужденное состояние, за счет чего осуществляется синтез органических соединений - фотосинтез. Понятие о ФАР ввел Л. А.
Иванов (1918), назвав ее «физиологически активной радиацией», развил и широко внедрил его А. А. Ничипорович (1956). Рядом исследований было показано, что в зависимости от высоты
Солнца и прозрачности атмосферы содержание ФАР в суммарной интегральной солнечной радиации, падающей на Землю, может меняться от 28 до 42 - 43%. в рассеянной радиации ФАР
содержится до 60%.
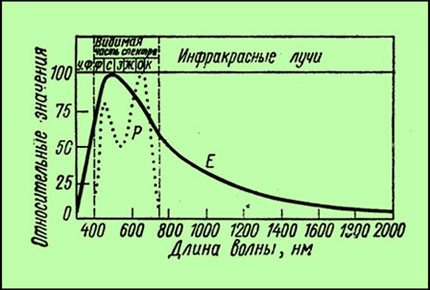
Если рассматривать распределение энергии общей солнечной радиации, падающей на Землю, то из хода кривой видно, что максимум энергии приходится на видимую часть света. Солнечный
свет сильно меняется при прохождении через атмосферу. Поэтому если на верхней границе атмосферы максимум энергии солнечного света отмечается при 470 нм, т. е. в синей части, то
на поверхности Земли при высоком летнем солнцестоянии максимум энергии уже приходится на желтую часть, а при низком положении солнца (на восходе или при закате) - на красную.
По действию энергию солнечных лучей, падающих на лист, можно разделить на 4 физиологические зоны:
1. Зона действия волн, длиной 300 - 520 нм. Эти волны поглощаются хлорофиллом, каротиноидами, протоплазмой, ферментами. Но все же большая часть их поглощается именно хлорофиллом.
2. В зоне 520 - 700 нм поглощение связано исключительно с хлорофиллом. Эти длины волн соответствуют оранжево-красным лучам, значение которых для фотосинтеза показал еще К. А.
Тимирязев. Такая радиация имеет решающее значение для всех физиологических процессов - фотосинтеза, развития, формообразования и т. д.
3. Зона 700 - 1050 нм - инфракрасные лучи, т. е. так называемая «абиотическая радиация», не играющая почти никакой биологической роли.
4. Зона свыше 1050 нм - дальняя инфракрасная радиация, мощный фактор теплового режима, хорошо поглощаемая цитоплазмой, водой и т. д.
Наиболее интенсивное поглощение радиации листом (первый, больший максимум) проявляется в оранжево-расных и красных лучах (600 - 680 нм) и второй (меньший) максимум поглощения -
в ультрафиолетовых лучах (300 - 520 нм); минимум поглощения приходится на желто-зеленые лучи (550 - 575 нм). Поглощение в инфракрасной части вызывает нагревание листа, но при
низких температурах эти лучи тоже частично поглощаются хлорофиллом и, видимо, могут быть полезными для фотосинтеза. Во всяком случае, было показано, что устранение инфракрасной
радиации приводит к частичному снижению продуктивности. Желто-зеленые лучи поглощаются листом слабее и не влияют на фотосинтез непосредственно, но они, видимо, являются источником
сильного светового возбуждения, регулируют световое раздражение и этим оказывают влияние на фотосинтез. Ультрафиолетовые лучи имеют губительное воздействие на живые организмы.
Так, при моментальном облучении ими слой воды глубиной 30см становится совершенно стерильным. Правда, в растение эти лучи проникают на небольшую глубину: под их воздействием
обычно отмирает эпидермис, а цитоплазма лежащих глубже клеток оказывается защищенной. Ультрафиолетовые лучи очень хорошо поглощаются антоцианом (anthocyan); возможно, этот пигмент
действует как экран, защищая растение (кстати, антоциана много у высокогорных растений). На хлорофилл ультрафиолетовые лучи почти не действуют, однако у этиолированных растений
под их влиянием он интенсивно образуется.
Растения используют не только прямой, но и рассеянный свет. Прямые солнечные лучи часто опасны для растений, ибо при их большой интенсивности разрушается цитоплазма и хлорофилл.
Рассеянный свет обычно усваивается полнее, и, кроме того, он выгоднее и по составу - в нем до 50 - 60% желто-красных лучей (ФАР), важных для фотосинтеза, а в прямом их бывает
всего 30 - 35%. Наконец, дневной свет состоит не только из прямого и рассеянного солнечного света, но к нему добавляется еще диффузный свет, отраженный от неба. Поскольку
хлорофилл лучше всего воспринимает лучи красной и синей частей спектра, то полнее используется свет, когда он слаб, например при низком стоянии солнца, при облачности и т.
Несколько хуже хлорофилл воспринимает свет при высоком положении Солнца, так как в это время энергетический минимум света лежит в желто-зеленой части спектра.
ПРЯМОЙ И РАССЕЯННЫЙ СВЕТ
При изучении светового режима очень важно различать воздействие прямого и рассеянного света. Известно, что атмосфера, особенно ее «переменные компоненты», такие, как водяной пар,
СО2 и молекулы других газов, очень сильно влияет на солнечную радиацию. Примерно 42% радиации, достигающей атмосферы, снова отражается и рассеивается в мировом пространстве, т. е.
альбедо равно 42%. Приблизительно 15% (из 58%) поглощается, абсорбируется атмосферой, следовательно, поверхности Земли достигает только 43% солнечной радиации (солнечной постоянной).
Свет, рассеянный облаками и прошедший через них, сильно обеднен коротковолновой ультрафиолетовой, сине-фиолетовой и инфракрасной радиациями. Отсюда следует, что в рассеянном
свете больше полезных для фотосинтеза оранжево-красных лучей, чем в прямом, т. е. прямой свет, падающий на лист, очень сильно отличается от рассеянного, называемого диффузным.
В ясные дни диффузный свет составляет 10 - 15% от общей радиации, а в пасмурные - все 100%. Однако соотношение между рассеянной и прямой солнечной радиацией, между интенсивностью
света и его спектральным составом крайне изменчиво и неодинаково в разных географических условиях, на разной высоте над уровнем моря, оно зависит от состояния атмосферы, рельефа,
характера растительности и т. д. Эти соотношения различны в разные часы дня, в разные сезоны вегетационного периода, в разные годы.
Поскольку интенсивность радиации сильно уменьшается при прохождении через толщу атмосферы, то в высокогорных альпийских областях, где слой атмосферы тоньше, она намного больше, чем
в низменностях. Уменьшается интенсивность радиации при низком стоянии Солнца над горизонтом. Так, при высоте Солнца 60° она составляет 916 Дж х м−2 х с−1;
при высоте 30° уменьшается до 699, а при высоте 5° падает до 272 Дж. м−2 х с−1. Надо иметь также в виду, что и сама высота солнцестояния меняется не
только в течение суток, но также по временам года и в зависимости от географической широты. Как правило, утром и вечером из-за низкого стояния Солнца отмечается больше рассеянного
света с преобладанием длинноволновой радиации, а в полдень преобладает прямой свет, богатый коротковолновой радиацией, однако измерения радиации обычно проводятся для горизонтальных
поверхностей. В природе огромное значение имеют экспозиция и наклон склонов. Изменение угла падения солнечных лучей сильно меняет и интенсивность радиации. Поэтому более заметны не
широтные, а топографические вариации силы света. Это особенно относится к высоким арктическим широтам, где различия в интенсивности южных и северных склонов значительнее, что во
многом объясняет и разницу в растительности этих склонов.
Рассеянный свет, составляющий в среднем 1/10 интенсивности прямых лучей, поглощается растением почти полностью, и коэффициент его использования намного больше. Поэтому в северных
широтах преобладающего рассеянного света вполне достаточно для интенсивного продуктивного фотосинтеза. Но здесь продуктивность ограничивается не светом, а низкими температурами.
Если, например, сравнить количество солнечной энергии, приходящейся в течение года на единицу площади, то окажется, что в Крыму оно в 5 раз больше, чем на Шпицбергене, и в 2 раза
больше, чем в Ленинграде. Но за 4 месяца вегетационного периода (с мая по август) эти различия сильно сглаживаются, особенно если учесть рассеянный свет, которого на севере больше.
Было, например, подсчитано, что с 1га на Шпицбергене (если иметь в виду только световое довольствие) можно бы собрать столько же продукции, сколько с 1га в средней полосе. Но из-за
недостатка тепла в Арктике значительная часть света оказывается недоиспользованной.
Во влажных тропиках водяные пары оказывают большое экранизирующее влияние на свет, поэтому там освещенность значительно снижается. В пустынях из-за, малого количества водяных паров
интенсивность света значительно выше, чем в гумидных областях, а это позволяет пустынным растениям сокращать транспирационную поверхность без особых последствий для ассимиляции. Но
обильный свет в пустынях также полностью не используется растениями из-за высоких температур и недостатка воды. Обычно отмечается, что при движении от экватора к полюсам поглощение
солнечной энергии атмосферой увеличивается, и ее поступает на поверхность почвы все меньше и меньше. Однако при движении к северу одновременно увеличивается и количество рассеянного
света. Поэтому в умеренном поясе летом суммарной солнечной энергии получается больше, чем на экваторе, да и содержание в ней губительных ультрафиолетовых лучей падает.
Кроме прямого и рассеянного (бокового) на открытых местах растения получают свет, отраженный от поверхности почвы (а зимой - от снега), т. е. нижний свет. Снежная поверхность отражает
в среднем до 30% (а чистый снег - до 80%) суммарного дневного света, причем спектральный состав отраженного света почти не меняется. Зеленый луговой травостой отражает 4% поступающего
на луга света, преимущественно длинноволновой части спектра. Особенно интенсивен нижний свет на южных склонах берегов рек, озер и морей, где в солнечный день он может составлять
35 - 85% от прямого. По-разному отражают и пропускают свет различные растения, поэтому световой режим в сообществах неодинаков.
ИЗМЕРЕНИЕ РАДИАЦИИ И ОСВЕЩЁННОСТИ
В климатологии величина общей радиации обычно измеряется актинометрами. Для целей экологии этого недостаточно. Обычные актинометры представляют собой два термометра, помещенные рядом
в один футляр, но резервуар одного из них выкрашен в черный цвет, а другого - в светлый. По разности температур с помощью переводных таблиц определяют радиацию в калориях на единицу
площади в единицу времени. Л. А. Иванов применил так называемый «фитоактинометр», который тоже имеет два термометра, но в одном из них чистый (прозрачный) толуол, а в другом раствор
хлорофилла в толуоле, который поглощает фотосинтетически активную радиацию. «Зеленый» термометр, нагревается сильнее, чем прозрачный, и разница в пока¬заниях термометров
пропорциональна поглощенной хлорофиллом энергии; по переводной таблице определяют радиацию в калориях на единицу поверхности в единицу времени.
Для количественного выражения различий в освещенности раньше пользовались приемами, основанными главным образом на скорости почернения фотобумаги. Этим методом много работал
австрийский ученый Визнер. Но данным методом можно исследовать фактически лишь ультрафиолетовую часть радиации, к которой чувствительна фотобумага. В последнее время пользуются
люксметрами, воспринимающей свет частью которых являются селеновые фотоэлементы. Шкала прибора показывает значения освещенности в люксах (лк). Датчик люксметра (селеновый фотоэлемент)
имеет пределы измерений от 380 до 710 нм с максимумом чувствительности при 550 нм. Соотношение между освещенностью в люксах и ФАР (энергетический эквивалент люкса), по расчетам Ю. Л.
Цельникер (1969), следующее (при высоте Солнца 40 - 500): 1 лк = 5,7 х 10−6 кал х см−2 х мин−1 (в границах длин волн 380 - 710 нм).
Такой эквивалент позволяет устранить искажающее влияние избирательной чувствительности люксметра на результаты измерений. Величина эквивалента сильно усреднена и может значительно
меняться при различных условиях. Однако при работе с люксметром надо учитывать и ошибки измерений, которые зависят от высоты солнца и температуры. Кроме того, чувствительность
люксметра снижается при высоких интенсивностях света (ошибка может достигать 12%).
СВЕТ И ЖИЗНЕННЫЕ ФУНКЦИИ РАСТЕНИЯ: СВЕТ и РОСТ
Всходы ели в еловом лесу, как правило, гибнут из-за недостатка света, так как в растении не накапливаются органические вещества, а запас пластических веществ в семенах слишком мал.
Света несколько больше под пологом березы, но лучше всего сеянцы ели развиваются на вырубках, где достаточно света и отсутствует корневая конкуренция.
Значительная освещенность ингибирует вытягивание междоузлий, что влияет на размеры растущего побега у многих древесных пород рост в высоту почти не меняется при интенсивности света
50 - 90% от полной освещенности. Однако в случае меньшей освещенности растения бывают низкорослыми. Ю.Л. Цельникер (1968) отмечал, что уменьшение освещенности влияет на интенсивность
фотосинтеза, а это приводит, в свою очередь, к снижению прироста у разных пород по-разному, а у теневыносливых растений в наименьшей степени. Так, прирост не наблюдается (т. е.
баланс органических веществ равен нулю) у березы и осины при интенсивности света 4,5% у клена, дуба и ясеня 3,0 %, у каштана конского и липы – при 2%.
При крайних затенениях (менее 0,5%) у многих древесных пород уже отмечаются заметные явления этиолированности, особенно у растений, имевших к моменту падения освещенности
достаточный для вытягивания побегов запас питательных веществ в семенах или в органах запаса (клубни). Если же такого запаса не было, то у растений явления этиоляции не развиваются
или же они гибнут от недостатка света. Влияние света на рост (как стимулирующее, так и через фотосинтез) во многом определяет успех в конкуренции за свет между растениями
в фитоценозе. Это особенно заметно в темном тропическом лесу, где деревья, вытягиваясь, выносят свои кроны выше затеняющего их полога. Наконец, рост довольно тесно связан с
фотопериодизмом, о чем будет сказано ниже.
ВЛИЯНИЕ СВЕТА НА НАПРАВЛЕНИЕ РОСТА
Многие растения, особенно световые, обладают свойством расти в сторону источника света, если их освещать с одной стороны. При повороте растения на 180° верхушка стебля опять
изогнется в сторону источника света. Это свойство называется фототропuзмом. У стеблей он положителен, т. е они растут в сторону источника света, у корней - отрицателен
(«уходят» от света). Наконец, у многих листьев, располагающихся перпендикулярно к направлению света, фототропизм «поперечный». Фототропические явления вызываются перераспределением
особых ростовых веществ - ауксинов, вырабатываемых верхушкой стебля и корня. Классические работы с этими веществами проделал М. Х. Чайлахян. Фототропическая реакция
подчиняется так называемому «закону количества раздражения», иначе говоря, важны не фактическая интенсивность света и не продолжительность освещения, а общее его количество, т. е.
произведение интенсивности на продолжительность.
Эффект фототропизма объясняется разницей в скорости роста на затененной и освещенной сторонах органа - большее торможение происходит на освещенной стороне, что и вызывает искривление
или поворот органа. При освещении листьев со всех сторон они располагаются обычно горизонтально, но у некоторых растений (эвкалипт, гладиолус, ирис, злаки) расположение листьев
приближается к вертикали. На своеобразное расположение листьев у «компасных растений» помимо температуры влияет, конечно, и свет - утренний и вечерний, что позволяет им избегать
полуденной переосвещенности. Экологическое значение фототропизма очевидно: для стеблей и листьев важно занять такое положение, при котором растение будет получать оптимальное
количество света.
Организмы, обладающие свободным движением (жгутиковые, пурпурные бактерии и др.), способны перемещаться по направлению к источнику света. Это фототаксис. Другим примером
фототаксиса можно назвать перемещение взвешенных в цитоплазме хлоропластов под влиянием света: в темноте хлоропласты располагаются более или менее равномерно; при слабом освещении
они перемещаются на освещенную стенку клетки, а при сильном, прямом, солнечном свете хлоропласты переходят на боковые стенки и свет падает на грани.
Фотонастии. Направление ростовых реакций зависит положения источника, т. е. от направления падающего света. Но иногда направление роста стеблей и листьев обусловливается только
строением реагирующего органа, хотя эта реакция вызвана освещением. Например, направление роста боковых побегов шалфея висит от интенсивности освещения: чем она сильнее, тем
горизонтальнее расположены боковые побеги, при этом освещение со всех сторон равномерное, т.е. фототропизма нет. Следовательно, здесь реакция вызвана светом, а не его направлением.
У хризантем боковой побег на свету растет вертикально, если же точку его роста прикрыть; то он занимает горизонтальное положение. Когда все растение находится в темноте, то основной,
главный, побег постепенно полегает, т. е. занимает тоже горизонтальное положение. Но как только растение попадает на свет, его стебли снова выпрямляются. У тех же хризантем при
выращивании в темноте листья растут вертикально, но в этом случае сильнее вытягиваются клетки нижней поверхности листа (гипонастия). На свету они переходя т в горизонтальное положение,
т. е. энергичнее растет верхняя сторона (эпинастия). Подобное же явление наблюдается у молодых растений фасоли. У подорожника среднего при очень высоких интенсивностях света листья
сильно изогнуты книзу (эпинастия), а при меньших интенсивностях света они стоят вертикально (гипонастия).
С влиянием света связаны и нuктuнастuческuе движения, например многие цветки и соцветия (одуванчик, мать-и-мачеха) открываются утром и снова закрываются к ночи. Некоторые
растения (мак) раскрывают свои цветки на рассвете, другие (лен) - раскрывают их на несколько часов позднее; у дурмана цветки открываются только вечером. Аналогичные явления отмечены
и у листьев. Правда, здесь помимо света, видимо, влияет и изменение тургора. Например, черешки фасоли принимают более вертикальное положение ночью, а пластинки в это время больше
наклоняются к земле, чем ночью. У томата листья, горизонтальные днем, ночью занимают более вертикальное положение; листочки клевера на ночь складываются кверху и вдоль средней жилки;
складываются при сильном освещении и листочки кислицы. Все эти движения стебля и листьев под влиянием света имеют определенное экологическое значение.
РЕПРОДУКЦИЯ
Было отмечено, что недостаточное освещение подавляет цветение, а иногда вообще задерживает растение в состоянии вегетации. Отсюда интересен следующий вывод: поскольку
пониженная интенсивность света благоприятствует развитию вегетативных органов за счет репродуктивных, то, следовательно, урожай, выращиваемый ради вегетативных частей, будет большим
при высоком проценте облачности. А для получения урожая плодов, зерна, семян необходимы яркий свет и ясная погода. Для морского, влажного климата, видимо, предпочтительнее положительные
отклонения от сезонной нормы освещенности, т. е. в сторону увеличения числа ясных дней. А для континентального климата, с его сухими ясными периодами, предпочтительнее отрицательные
отклонения от нормальной сезонной освещенности, т. е. в сторону увеличения облачности.
ТРАНСПИРАЦИЯ
Вредный эффект высокой интенсивности света сказывается в том, что повышается транспирация. Свет стимулирует открывание устьиц, повышает чувствительность мембран. Краткосрочное яркое
освещение действует на устьица разных древесных пород по-разному. Например, устьица теневыносливого бука открываются быстрее, чем устьица светолюбивого тюльпанного дерева, виды со
«средней» теневыносливостью (клен, дуб) проявляют промежуточную реакцию. Кроме того, устьица на верхней и нижней сторонах листа реагируют по-разному: у табака закрывание устьиц верхней
поверхности листа вызывалось освещенностью в 10 раз большей, чем это требовалось для устьиц нижней поверхности. В случае иссушения теневые растения под воздействием света закрывают
устьица быстрее, чем световые. Следовательно, устьица листьев растений из затененных местообитаний реагируют на повышение освещенности быстрее, и их реакция меньше ограничена низкими
освещенностями, чем у растений открытых местообитаний. Это дает им возможность четче регулировать фотосинтез при значительных колебаниях освещенности.
|
|
|
|
График распределение энергии общей, падающей на Землю, радиации (Е) и относительная интенсивность фотосинтеза (Р) в лучах различной длинны волн; УФ – ультрафиолет; части спектра:
Ф – фиолетовая; С – синяя; З – зелёная; Ж – желтая; О – оранжевая; К – красная.
|
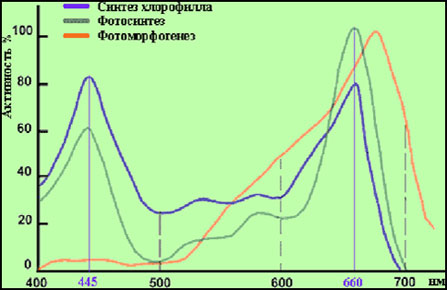
График активности фотобиологических процессов: наиболее интенсивный пик поглощения приходится на желто-красную (600-750 нм) область, меньший – на сине-фиолетовую
(400-500 нм), так же виден "провал" в области зеленой (500-600 нм) части спектра.
|
|
Copyright © 2008 - 2017 Emil Morozov. All Rights Reserved
Копирование, использование и размещение материалов данного сайта на других
ресурсах, без согласия автора, запрещено. |

E-mail: emil-morozov@yandex.ru |
| |